
This series of posts are inspired by two questions that Jarrett Byrnes asked:
- Given the extinction of E predators out of N, what is the probability that a prey species will still have at least one predator remaining?
- Given E out of N, what then is the probability that all prey species will have at least one predator remaining?
As Jarrett and I have been discovering, these are actually quite difficult questions to answer in a general manner, i.e. for all topologies of a certain size!
Say we have a two trophic level food web with N predators, what is the probability that a prey species has at least one predator remaining after the extinction of E predators? The solution provided here depends on having the out-degree of prey species, and finding the probability that all predators of a prey species become extinct as a result of E. Say that the out-degree of the prey species is 
which reduces to
The probability then of a prey species having at least one prey is 1 minus the above
that is, the sum of the probabilities of having 1 predator, 2 predators, etc.


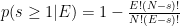
Example
Let the adjacency matrix of a food web be
where predators are rows and prey are columns. Our prey out-degree set is therefore {3, 2}. For E=1, both prey will have at least one predator since their out-degrees both exceed 1. For E=2, the possible resulting topologies are
For s=2
This is correct since our prey species of out-degree 2 (second column of A) has at least one predator in two of our three post-extinction topologies. The probability should be zero for s=3 (since E<s). If we add a third prey species, with s=1, making
then for E=1, the post-extinction topologies are
The probability that this third species has at least one prey is also 2/3.




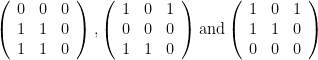

A further example
So far so good, right? Well, Jarrett posed this example,
Notice that we now have two prey of out-degree 2. For E=1, the post-extinction topologies are
Applying the above formula yields
which is correct, since two of the three topologies maintain at least one predator for each prey. When E=2, the topologies become
Obviously, 
What went wrong?! The answer points to just how devilish the questions are, and how deceptive! There are two species of out-degree 2 (s=2) in the food web, hence the second term in the formula is squared (see above). BUT, the predator-prey topologies of the species are different, meaning that simple hypergeometric counting cannot work. We literally must list and examine all the post-extinction topologies, but this is prohibitively impractical for food webs and networks of even modest size (a dozen species). So there we stand. We currently have a partial solution, and I will explore the difficulty and the partial solution in the next post.



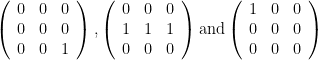
![p(s\geq 1\vert E=1) = \left [ 1 - \frac{2!(3-1)!}{3!(2-1)!}\right ] \left [ 1 - \frac{2!(3-2)!}{3!(2-2)!}\right ]^{2} = \frac{4}{27}](https://s0.wp.com/latex.php?latex=p%28s%5Cgeq+1%5Cvert+E%3D1%29+%3D+%5Cleft+%5B+1+-+%5Cfrac%7B2%21%283-1%29%21%7D%7B3%21%282-1%29%21%7D%5Cright+%5D+%5Cleft+%5B+1+-+%5Cfrac%7B2%21%283-2%29%21%7D%7B3%21%282-2%29%21%7D%5Cright+%5D%5E%7B2%7D+%3D+%5Cfrac%7B4%7D%7B27%7D&bg=ffffff&fg=000&s=0&c=20201002)

Ha! FYI, I’m starting a series on all of this stuff here
Already spotted it dude. I’ll point my next post to your blog. I also have some ideas on reducing the Inclusion Exclusion problem that I’ll get to whenever I get to the next post.